Signal Theory Overview
Towards a
Unified Theory of Consciousness & Psychedelic Action
Version 1.04 : 1.13.2006
By James
Kent
Summary
Signal Theory presents a new
model for objectively measuring expanded states of consciousness based on
neural firing rate, synchrony of neural spiking, and intensity of signal
feedback recursion occurring within the sensory processing circuits of
neocortex. Signal Theory also proposes methodologies for mathematically
modeling the action of psychedelic 5-HT2A receptor
agonists in the production of diminished, amplified, and standing sensory
feedback loops in simple neural circuits. Using the basic tenets of Signal
Theory, we can define an empirical model of perceptual action in which both
normal waking consciousness and expanded psychedelic consciousness can be
defined. This document is intended to be a brief overview of the basics of
Signal Theory, and is for general public review as well as scientific peer
review in the hopes of moving towards a more refined model of consciousness and
psychedelic action in the human brain.
Foundations of Consciousness
To understand how Signal
Theory can predict specific perceptual alterations to normal waking
consciousness, one must understand how the brain parses sensory data to create
the moment-to-moment reality we experience as consciousness. There are many definitions of consciousness, but for
the purposes of this discussion, consciousness is defined as the ongoing process of the self
contextualizing external stimulus. Consciousness is not an end state of
being, it is an ongoing feedback process between perception and
contextualization; each experience is perceived, parsed into meaningful
context, and integrated into memory to provide context for the next experience,
ongoing, ad-infinitum, constantly updating through time to create the cohesive
narrative identity that we refer to as the self.
The three necessary components of waking consciousness are perception (external
stimulus), contextualization (internal processing), and memory (data storage
and recall). In the human brain, perception is handled by feedback interaction
between the sensory organs, the thalamus, the amygdala, and the various
advanced sensory processing areas of the neocortex. Contextualization of this
sensory data is handled by feedback interaction between sensory processing
cortices (external data resolution); the executive areas of the pre-frontal
cortex (the PFC, or working memory); and the long-term-memory (LTM) functions
of the rhinal- hippocampal complex in the medial temporal lobe
(pattern-matching and associative memory). And though memory is stored
non-locally (holistically) in associative synaptic connections all over the
cortex, LTM compression and recall is handled by the rhinal-hippocampal complex
in the medial temporal lobe.
The fourth component of human
consciousness is, of course, emotional response, which is arguably
not-essential to consciousness per se,
but is nonetheless integral in all discussions of
human consciousness. In a circuit model of the human brain, the amygdala works
in tandem with the sensory processing cortices to provide instinctive,
sub-rational emotional responses to sensory data, particularly data relating to
fear, panic, survival, and mating. The amygdala also has direct feedback connections
with the sensory cortices, the PFC, and the LTM structures in the temporal
lobe, and is tightly wired into the intensity of both perception and memory
storage and recall. These four components of the human brain – perception,
contextualization, memory, and emotional response – are the foundations of the
Signal Theory of Consciousness as detailed in this text.
Introducing Signal Theory
Signal Theory is the name I
have coined for a new method of defining and measuring states of consciousness,
particularly when explaining and predicting the mind-expanding powers of psychedelic
chemicals on the brain’s perceptual processing capacity. Signal Theory is
derived from Cognitive Theory, a modern school of neuroscience that seeks to
derive the functioning of the brain by identifying and following the active
neural pathways where specific sensations, thoughts, and cognitive processes
arise. Via modern scanning techniques, lesion studies, and pharmacological
studies, cognitive scientists have been able to pinpoint areas of the brain
responsible for specific functions of consciousness, and have successfully
demonstrated how these specific areas of the brain all link up in networked
circuits to process raw sensory data into what we perceive of as consciousness
from moment to moment.
Signal Theory takes the
cognitive model one step further, and seeks to define consciousness in terms of
the flow of sensory signal through
neural processing circuitry; flow that can be measured via neural firing rate,
feedback parity within recursive circuitry, and synchrony of neural spike
timing between parallel sensory processing circuits. In other words,
consciousness is not the hardware itself (neural circuitry),
it is the flow of electrical current which passes through that neural circuitry
that gives us thought, mood, and sensation. This flow of consciousness alters
subtly yet consistently from instant to instant, giving us a near-real-time
picture of reality as it unfolds before us. By defining consciousness in terms
of signal intensity and feedback recursion within parallel sensory-processing
circuits, Signal Theory can empirically demonstrate how the normal functioning
of the human mind can be turned up or down; filtered; distorted; gated;
delayed; and looped to create an infinite array of classic psychedelic
perceptual effects simply by introducing the proper chemical catalyst (such as
a psychedelic partial 5-HT2A agonist) into the neural network.
The Role of Feedback Recursion in Analytical Neural Circuitry
Since consciousness is an
ongoing process – literally a flow of electrochemical charge through the brain
– it can be assumed that this process has a fundamental form and properties
that can be accurately measured and logically predicted. We already know much
about how signal passes through the brain, and the various neural pathways
signal takes as it is processed into conscious thought. What is interesting
about sensory signal is that it diverges on many different pathways through
various different specialized circuits before finally emerging holistically and
fully integrated into what we perceive as real-time consciousness. The
processes of maintaining spatial and temporal awareness, identifying incoming
data, and maintaining contextual identity all are ongoing and cyclical; which
means that neural networks
rely heavily on real-time feedback loops between areas of the
sensory cortices, areas of working memory, and areas of LTM. Real-time feedback
occurs in the thalamus to screen out unimportant noise; in the sensory cortex
to enhance data resolution; in the pre-frontal cortex to maintain holistic
contextual awareness; and in the medial temporal lobe to ensure robust signal
comprehension and recall. In other words, incoming sensory signal is routinely
processed multiple times by various layers of the brain, and then is
re-processed and double-checked by working memory to make sure it is accurate
before being worked over (yet again) in your sleep before attaining full
integration into LTM. The process of data perception, analysis, and memory
integration relies heavily on signal recursion through the very same
neural circuits over and over again to both ensure data fidelity as well as facilitate neural plasticity
and the creation and reinforcement of new synaptic bonds.
Signal feedback occurs at
many places and on many levels in the brain. Any particular piece of data may be passed
through working memory multiple times until it is fully analyzed and “released”
in order to allow new stimulus to update the process of waking consciousness.
During this “mulling time” the human subject often drifts into a kind of
distracted reverie, much like daydreaming, where obsessive rumination or
analysis of a specific data set takes over all active attention. This
rumination may take a split second or many minutes depending on context, but if
you have ever paused to make sure you understood a question correctly before
answering, then you understand how a few moments of re-analysis and subsequent
re-parsing of the same data set may yield new insights with each successive
pass. Within the tenets of Signal Theory, this process of cyclical rumination
is referred to as a recursive feedback
circuit, a circuit which can be defined in terms of both structure (neural
wiring) as well as recursion intensity,
or the rate at which signal makes a complete pass through the entire analytical
circuit before starting all over again.
Re-tuning the Cascade of Consciousness
Emerging from the bottom-up,
sensation starts at the skin, passes up the brainstem, and reaches the cortex
where it diverges into a parallel network of specialized analytic feedback
filters. Much like a fountain, sensory signal gushes upward through fat neural
pipes and then cascades into a shower of parallel logical circuits -- circuits
literally meaning “circles” or “loops” of analytical processing. This is what I
refer to as the cascade of consciousness, which is much like a standing wave
formation that can analyze and retain any number of contextual cues almost
indefinitely. This cascade represents the flow of consciousness through
the brain, and can be measured in many ways using many different high-tech
scanning devices.
In our normal waking state,
we take this flow of consciousness for granted, and can assume that in most
healthy individuals that the flow of signal through our neural processing
circuitry is tuned
and running well enough to keep us upright and functional. However, what
happens to this delicate cascade of consciousness when we tweak the flow, upset
the filters, re-modulate the recursion, and let the
feedback run wild? The fundamental assumption of Signal Theory is that an
increase in recursion intensity within the analytical feedback circuits in the
cortex would necessarily lead to classic psychedelic perceptual results, and
thus classic psychedelic molecules, such as tryptamine 5-HT2A
agonists, are prime suspects for facilitating the formation of standing neural
feedback recursion in at least one or more layers of sensory processing
circuitry.
Signal Theory Presumptions
1. Psychedelic 5-HT2A agonists act as neural feedback recursion promoters
The primary presumption of
Signal Theory is that psychedelics, though targeted neural excitation, act as
promoters for the intensity of feedback recursion occurring within various
layers of neural circuitry. This feedback promotion may be stimulated via
direct action at the post-synaptic receptor or via secondary action in the form
of asynchronous signal leakage from the pre-synaptic axon terminal. The most
likely target for this action is in the layer V pyramid cells of the sensory
cortices, where 5-HT2A receptors have the greatest density.
2. Intensity of
Feedback Recursion = Intensity of Psychedelic Experience
A secondary presumption of
Signal Theory is that recursion intensity within the iterative analytical processes
of the sensory cortices is the primary
source of the distinct sensory amplification, perceptual distortions, standing
hallucinations, and expanded states of consciousness perceived in the
psychedelic state. Feedback recursion within states of consciousness can be
measured in terms of zero-gain circuits (normal waking consciousness),
amplified-gain circuits (expanded, psychotic, or psychedelic consciousness), or
diminished-gain circuits (inhibited, dissociative, or sedated consciousness).
Within the realm of psychedelic action, feedback recursion may be modulated
upward or downward over the duration of any given psychedelic trip, based on
dose taken, external sensory input, and direct user biofeedback.
3. There are Optimal
Rates of Circuit Recursion and Neural Spike Synchrony
A third presumption of Signal
Theory is that there are optimal rates of recursion and circuit synchrony where
specific states of consciousness spontaneously manifest. Obviously, normal
waking consciousness is a delicately tuned state, and any change, interruption,
or perturbation of normal neural firing patterns would necessarily create a
corollary change in the subjective state of consciousness. While it is well
known that pharmaceuticals can be used to modulate neural firing patterns,
there has been very little research into the various non-ordinary states of
expanded consciousness generated by psychedelic drugs. Given that there are
many specific psychedelic states that appear to be well beyond the normal range
of human consciousness, one would expect there to be precise biophysical
benchmarks where these states of consciousness emerge. While much attention has
been paid to psychedelic effect on the action potentials of individual neurons,
Signal Theory suggests that neural firing rate is only one of many factors in
measuring psychedelic action. Instead, Signal Theory predicts that psychedelic
action is caused by an overall gain in the intensity of signal feedback recursion,
and subsequent synchrony of neural spiking occurring within cortical circuitry
over the entire duration of pharmacological affect.
Much attention is also paid
to the ‘peak’ of psychedelic experience, where neural processing appears to take a ‘quantum
leap’ beyond normal cognitive functioning, perhaps suggesting a
holographic comprehension of reality instead of the normal flat
representation we expect. Signal Theory predicts that this ‘peak’ state of
psychedelic action can be precisely measured and mathematically modeled against
states of normal waking consciousness to empirically demonstrate how amplified
feedback recursion and spike synchrony within neural circuits directly affects
perception and analytical capacity. Conversely,
one would assume that over the course of a psychedelic session that there might
be transitional, re-modulatory states that lack synchrony, create confusion and
dissonance, and severely distort or interrupt cohesive neural processing. As a
practical example of using biofeedback create ‘optimal’ resonant firing patterns,
one only need think of a shaman chanting, drumming, singing, and using other
rituals that serve to modulate synchrony in neural firing patterns. Within the
framework of Signal Theory, the shaman acts as a resonant biofeedback driver in
the creation of optimized waves of recursion in the participant’s neural
circuitry, thus allowing all participants within the circle to “tune in” to the
same level of consciousness where the shamanic state spontaneously manifests.
If these three fundamental presumptions
of Signal Theory prove to be true, then it will be possible to derive a unified
model for describing the entire range of waking and expanded states of consciousness
in a way that is empirically demonstrable.
A Schematic Description of Signal Theory
To illustrate the
fundamentals of Signal Theory, it is helpful to view a basic schematic of
sensory processing pathways within the brain. For simplicity, I have chosen to
illustrate the audio pathway, though these schematics can be adapted to the
visual and somatic pathways as well. The following illustrations represent two
different representations of the same audio-processing pathways. These
schematics are accurate, but over-simplified to demonstrate the levels of the
signal-processing workflow where feedback loops are likely to occur.
Figure 1: Crude Cognitive
Workflow of Auditory Signal Processing
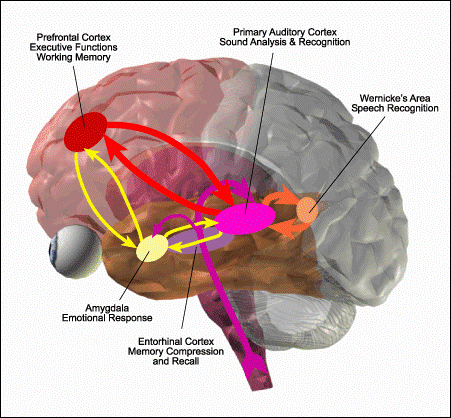
Figure 2: Flat
Schematic Workflow of Auditory Processing Pathway
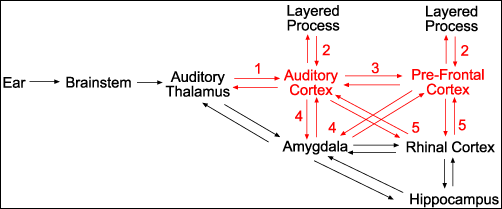
In Figure 2, sensory signal flow originates from raw sense data
hitting the ear (at left) and continues upward through the brain towards our
waking image of consciousness, which emerges at the far right in working memory
in the pre-frontal cortex (PFC) and rhinal-hippocampal long-term-memory (LTM)
systems in the medial temporal lobe. Along this signal processing pathway there
are many circuits which use feedback to control upstream signal flow,
illustrated by the double-arrow connections. These feedback circuits allow both
feed-forward and feed-back excitation and filtering of incoming signal
processing. The cortical areas with the highest densities of 5-HT2A receptors (such as the layer V pyramid cells in the
audio and pre-frontal cortices) are shown in red,
and the feedback circuits in red are those most
likely to be excited in the presence of a 5-HT2A agonist. These red feedback circuits have been
numbered by type, with the description of each type of circuit detailed below:
- Thalamocortical
feedback circuit: This circuit
connects the sensory thalamus to the sensory cortex. An increase in
recursion intensity in this circuit can lead to amplification of signal
strength, signal distortion, and temporal delay of signal data.
- Intra-cortical
feedback circuit: Processing in
the sensory cortex is done in layers of interconnected neurons which are
responsible for assembling fragmentary snippets of sensory data into
holistic representations of reality. An increase in recursion intensity in
these circuits can lead to increased detail resolution;
hyper-articulation of detail; signal noise and distortion; phantasmagoria;
and hallucinations.
- Inter-cortical
feedback circuit: Signal from
divergent sensory processing pathways converges on the pre-frontal cortex to
create the holistic, multi-modal sensory awareness we perceive as waking
consciousness. Constant feedback between the PFC and the sensory cortices
of the brain is essential to maintaining fidelity of signal and synchrony
of multi-modal sensory convergence. An increase in recursion intensity in
this circuit can lead to extreme temporal and perceptual distortions,
including sensory flanging, phasing, and echoes; recursive thought loops
and obsessive ideation; frame delay; moments replayed over and over in the
head; disappearance of time; and loss of multi-modal sensory cohesion.
- Amygdalo-cortical
feedback circuit: The amygdala
performs signal processing in networked parallel to the sensory and
pre-frontal cortices, monitoring sensory signal for potentially dangerous
stimulus. The amygdala regulates the body’s instinctive fear and panic
response, and is in constant feedback with all layers of sensory
processing to ensure robust signal fidelity and rational override in the
instance of false panic alarms. An increase in recursion intensity in this
circuit can lead to anxiety, paranoia, and panic.
- Rhino-Cortical
feedback circuit: The rhinal
cortex in the medial temporal lobe is sometimes referred to as the
transitional memory cortex, where information from various sensory
processing areas of the brain converge for
multi-modal memory compression in the hippocampus. These circuits are
essential for accurate long-term memory storage and recall. An increase in
recursion intensity in this circuit can lead to profound memory
imprinting, spontaneous memory recall; memory distortion; false memories;
temporary disruption of LTM storage (missing time, or blank spots) as well
as. Spontaneous activity in the medial-temporal is also known to cause
experiences which are mystical in nature, and patients with temporal-lobe
epilepsy often hear voices and have messianic inclinations, indicating
that recursion intensity in this circuit could potentially lead to a
variety of mystical states.
Although this is admittedly a
crude model of sensory signal processing in the brain, it does demonstrate the
basic tenets of Signal Theory and provides a working model in which accurate
predictions of sensory processing and subjective experience can be made in
terms of applied dosage and intensity of induced signal recursion.
Signal Theory and Cranial Blood Flow
While Signal Theory defines a
method of action for amplifying and distorting the normal functioning of the
human brain via the use of chemically mediated neural gates and feedback loops,
it does not totally explain why the same psychedelic drug may target a
different aspect of the mind or personality at different times in different
trips (or sometimes within the same trip). In other words, since psychedelics
are pharmacologically active in many parts of the brain, why would one LSD trip
be more visual when another may be more emotional? In order to explain this
targeted localization of psychedelic effect, I have come up with a model of
cranial blood flow in which the drug
follows the signal. In short, if a specific set of neural circuits becomes
activated, then cranial blood must flow to that area to feed the active circuits oxygen and glucose. If the psychedelic molecule
also moves through the bloodstream, the psychedelic molecule will follow in the
blood to the most active brain areas, and become pharmacologically active
wherever the brain is most active itself. Thus, psychedelic molecules can act
as non-specific amplifiers of any aspect of the user’s psyche, an effect which
is wholly targeted by the flow of signal, blood, and intent through the
individual subject’s mind.
Signal Theory Predictions of Experiential Psychedelic Phenomena
The Signal Theory model of
psychedelic action has been reverse-engineered from observing and cataloging
the subjective results of psychedelic action, then attempting to find a
theoretical cognitive model that could accurately predict the extreme range of
experiential results. With that in mind, it would seem that I am simply
re-stating what is already known about psychedelics when I say that Signal
Theory makes “predictions” about the phenomenological results of psychedelics.
Nevertheless, it is helpful to follow the theory back to action in order see if
the predictions indicated by Signal Theory indeed make sense.
Low Dose:
Signal Theory predicts that at a low dose of psychedelics, moderate signal
recursion in the layers of the sensory cortices would cause sensory signal to
become sharper; the resolution of detail would begin to stand out; and thought
in the rational forebrain would arise faster, sharper, and with heightened
intensity.
Moderate Dose:
Signal Theory would predict an overall increase in signal intensity in the
thalamocortical, amygdalo-cortical, and inter-cortical feedback circuits,
causing brief sensory echoes as well as hyper-articulation of all sensory
detail. At this point perceptual distortions become more acute, thinking
becomes more obsessive, anxiety and paranoia becomes more acute.
High Dose:
Signal Theory would predict increased intensity in inter-cortical and
rhino-cortical feedback circuits, creating sensory echoes which emerge and
swirl inward on themselves with diminishing intensity. At this point fully
articulated hallucinatory constructs begin to form with very limited temporal
cohesion; thinking becomes erratic; there is an increase in spontaneous memory
recall; there is an increase in temporal distortions; and multi-modal cohesion
of external stimulus begins to unravel.
Extremely High Dose: Signal Theory would predict standing recursion waves throughout all
sensory processing areas of the brain, allowing for the formation of fully
articulated hallucinatory constructs with high temporal cohesion. At this
point, the rational forebrain is overcome by information pouring in through
self-sustaining sensory feedback loops, and thought emerges holographically
into consciousness as fully-articulated multi-modal synesthesia.
Experiential Validation of Signal Theory
While the biomechanical
fundamentals of Signal Theory are still being explored, the basic model itself
can provide a great deal of insight into what the subjective consciousness may
be capable of experiencing under the influence of psychedelics. Using the basic
dynamics of Signal Theory, the subject can attempt to objectively parse what is
happening inside the head during the psychedelic trip, and even attempt to
apply the theory to targeted amplification or expansion of specific functions
of the mind. If a basic understanding of Signal Theory helps the subject
maintain a conscious level of control and mastery over the psychedelic state,
then the model is experientially valid, and could have a broad application in both personal, professional, and therapeutic use of
psychedelics. While experiential validation is a long way from empirical proof
of the validity of Signal Theory, it does at least provide a workable model for
thinking of the psychedelic state without having to invoke either psychosis or
spirits to explain the unique experiential results.
Empirical Validation of Signal Theory
Signal Theory may be tested
and validated in a number of objectively measurable ways. The first step would
be to locate the primary areas of action where sensory signal feedback and
iterative neural processes are most likely to occur. As the diagrammatic model
of Signal Theory shows, the primary areas to watch for signal recursion and
feedback excitation would be between the sensory thalamus, the layer V pyramid
cells of the sensory processing cortices, and the projections which lead from
the sensory cortices to the multi-modal convergence points in the pre-frontal
lobe and transitional memory cortex (and back). In actuality, there may be
multiple layers of signal recursion and feedback amplification occurring
between many levels of the brain at all once, but charting the densities of 5HT2A receptors throughout the cortex and following their
afferent feedback connections seems like the logical starting place to look for
the kind of iterative feedback circuits Signal Theory predicts.
Secondly, once an area (or
areas) of the brain are identified as potential pathways for sensory signal
recursion, scanning and monitoring methods must be developed to measure neural
activity and blood flow to these areas both in baseline settings and during
targeted psychedelic states. Signal Theory predicts that specific areas of the
brain will increase in neural firing rate to correspond to the intensity of
specific psychedelic phenomena, and that the synchrony of neural firing will
remain constant between all active brain areas where standing feedback
recursion is occurring. Though the impact of signal recursion may theoretically
be measured on one or more cranial scanning devices, getting precise rates of
recursion from animal subjects would most likely require the implantation of
electrodes into various areas of the brain to monitor inter-cortical signal
recursion and intensity in a more precise manner.
Mathematical Exploration of Signal Theory
Though I am presently not
familiar with any mathematical models to describe the intensity of signal
recursion within neural feedback circuitry, I must also confess that my overall
knowledge of computational neurobiology is limited, and that I have no capacity
to adequately test any mathematical models I may come up with or come across on
my own. But moving towards a mathematical model of neural signal recursion, a
few primary factors would need to be adequately articulated as a starting
point. To simplify this process, I present a crude two-neuron schematic of an
iterative feedback circuit for deconstruction below:
Figure
3. Simple Iterative
Neural Feedback Circuit
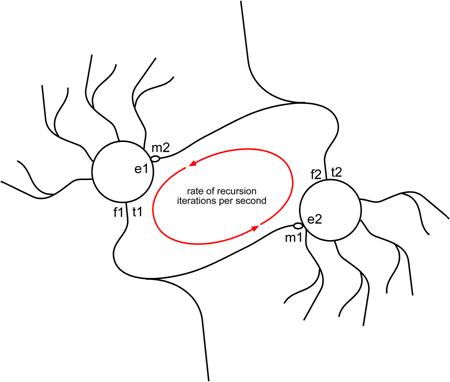
In Figure 3.,
two neurons create a simple iterative feedback circuit. The cell-body on the
left is neuron 1 (n1), the cell-body on the right is neuron 2 (n2).
Feed-forward signal from (n1) passes to (n2); feedback signal passes from (n2)
to (n1). The primary factors that can be measured or deduced from this model
are as follows:
Recursion Intensity Primary Factors
(f1) firing rate : feed-forward firing rate (frequency, in hz) of signaling neuron.
(t1) firing time: time of
(n1) action potential (start at 0 for calculating circuit synchrony).
(f2) feedback rate: firing
rate (frequency, in hz) of
post-synaptic feedback neuron.
(t2) firing time: time of
(n2) action potential (0+ milliseconds for calculating circuit synchrony).
(e1) and (e2) excitation:
ratio which measures relative excitation of neural membrane at post-synaptic
receptor site. 1e would be normal resting state (neutral), 1.2e would be a
slightly excited state, .8e would be a slightly
inhibited state. This factor expresses primary pharmacological interaction at
the receptor.
(m1) and (m2) modifier: ratio
which measures relative asynchronous transfer rate of signaling transmitters
leaked from the pre-synaptic axon terminal. 1m would be normal 1:1 synaptic
firing to signal transmission. 1.2m would indicate slight asynchronous leakage
of transmitter at pre-synaptic terminal. This factor expresses secondary
pharmacological interaction at pre-synaptic axon terminal.
(p) feedback
parity: ratio of feed-forward output frequency to feed-back input frequency.
Perfect parity = 1; slightly diminished parity = .8; slightly amplified parity
= 1.2. Parity would be a good first indicator in calculating feedback recursion
intensity and iterative signal decay.
(ips) iterations per second : the number of iterations
signal makes through the neural circuit per second, expressed in hertz. The
more hz, the greater the
rate of recursion.
(rr) rate of recursion: time in milliseconds to
complete a signal feedback circuit; another means of expressing iterations per
second.
(s) synchrony
: delay, in milliseconds, between firing of successive neurons in a
circuit. Perfect synchrony would be 0, in which all neurons in the circuit were
firing at precisely the same moment to create a standing wave of signal
recursion.
(rs)
resonance : a factor which modifies synchrony.
Resonance can be calculated using (f) and (t) to mark sympathetic frequency
amplifiers between neurons in synchronized circuits.
(d) decay:
time, in milliseconds, in which recursive feedback signal decays in a given
iterative circuit. Can be observed and averaged over time, or may be generally
predicted using feedback parity (p), excitation (e), synchrony (s), and
asynchronous modifiers (m) as indicators of potential signal decay or
amplification over successive iterations.
Signal Decay and Recursion Intensity
Normal sensory signal decay
is fairly quick in terms of milliseconds; images fade from our mind very
rapidly when we close our eyes. Signal Theory predicts that an increase in
recursion intensity and circuit synchrony would also increase the length of
time it takes that signal to decay over time (thus trails, feedback loops,
after-images, etc). In other words, signal decay should increase in direct
proportion to parity, synchrony and rate of recursion, and be modified
incrementally by the pharmacological factors (m) and (e). And though it would
be premature to try and model those variables without any experimental data to
plug in and test, it seems clear that parity, circuit excitation, synchrony,
and signal decay would be the major variables to look at when attempting to
measure the relative recursion intensity of a particular neural circuit over
time.
Notes on Calculating Synchrony, Resonance, and Standing Recursion
Synchrony and resonance may
be the most difficult factors to calculate within complex neural circuits in
living subjects. In a simple two-neuron circuit, s = (t2-t1), and if both
neurons are firing rhythmically at the same moment, synchrony becomes a perfect
0. If synchrony becomes a perfect 0 and feedback parity reaches a perfect 1,
then a perfect standing recursion wave has been achieved. It is unknown if
human neural circuitry can actually achieve a perfect standing recursion wave,
but we know it can certainly come very close. If synchrony is near 0 and parity
is .8, you have what would be measurable feedback recursion with predictable
signal decay. If synchrony is near 0 and parity is at 1.2, you have an
amplified recursion wave that grows in intensity with each
iteration. However, there are many ways that neurons in a complex
circuit can fire in resonant synchrony, meaning a standing wave of recursive
signal is generated not from perfect synchrony, but from neurons firing in
sympathetic rhythmic patterns that actually drive signal strength. Also, we are talking
about modeling the action of thousands of neurons all firing within a
hundredth-of-a-second of each other, so closing the gap on synchrony and
resonance is literally a kind of temporal hairsplitting. In truth, identifying
and modeling all the different rhythms, syncopations, and cohesive resonant and
dissonant patterns that arise in complex standing neural iterative processes
may become a scientific sub-field unto itself. The technology for scanning and
measuring these distinct neural firing patterns is among us today, but the
basic questions in this field still remain unanswered. Perhaps computer models
can be generated to model the kind of output intensity Signal Theory predicts,
but until a more accurate method for calculating synchrony in complex neural
circuits is devised, we can only guess at how much of a factor synchrony and
resonance are in generating standing signal recursion in neural structures.
Developing a Mathematical Model
What I offer here is only a
brief mathematical discussion, which is the first steps towards exploring
Signal Theory and developing a model that can be measured and predicted not
only in experiential results, but in tests on electrical current running
through slices of neural tissue in vitro.
However, if one would like to examine examples of mathematical models for
calculating synchrony and spike timing in complex neural networks, please
follow the relevant references presented at the end of this overview.
Signal Theory Questions
The Role of Iterative Feedback Circuits in the Production of
Consciousness
What are the functions of
iterative neural feedback networks in normal sensory signal processing? Signal
Theory would predict that they are instrumental in resolving fine detail from
ambiguous data sets, and refining crude sensation into articulated thought. In
other words, Signal Theory predicts that iterative neural analysis enhances signal
resolution and articulation of detail in the same way that multiple optical
scans of a photograph can produce far greater detail resolution than a single
scan. In other words, the detail is in
the recursion.
Dopamine as Promoter
of Neural Firing Synchrony in Iterative Circuits
What is dopamine’s role in
the formation standing recursion waves? Given dopamine’s role in ADD,
psychosis, and mediating fine motor control, evidence suggests that dopamine
may be essential to modulating neural firing synchrony in the formation of
standing resonant patterns. Given this assumption, Signal Theory predicts that
dopamine is also essential in modulating and fine-tuning resonant synchrony in
standing waves of sensory feedback recursion in the psychedelic state. Too little
dopamine would lead to the inability to focus the mind to form standing
recursion waves; too much dopamine would lead to the spontaneous eruption of
standing recursion waves with high temporal cohesion. Since fine motor control,
concentration, and multi-modal sensory convergence all rely on recursive
feedback circuits to fine-tune neural synchrony, the basic tenets of the dopamine model of
psychosis, Parkinson’s, and ADD dovetails precisely with the basic tenets of
Signal Theory.
Optimal Levels of
Recursion Intensity for Expanded Consciousness
Signal Theory predicts that
there are optimized levels of recursion and circuit synchrony for both normal
consciousness as well as various states of expanded psychedelic consciousness.
Can these optimal recursion rates be objectively measured, mapped, and
predicted? Can these states be spontaneously stimulated via focused biofeedback
or application of targeted transcortical stimulation? This is the big question Signal Theory poses.
Accurately identifying and predicting the optimal recursion factors that lead
to specific expanded states of consciousness is the ultimate goal of Signal
Theory, a goal that will not be achieved until further in-vitro an in-vivo
experimental testing can be done.
Signal Theory, Neural
Plasticity, and PHPD
Can recursive feedback
networks activated over time form lasting recursion pathways which persist
after the psychedelic molecule has been metabolized? Tenets of neural
plasticity would indicate that the longer and more frequently recursion
intensity is applied to any given circuit, the more likely it is that the
circuit will fall into standing signal recursion under normal conditions. The
role of iterative processing and circuit synchrony in neural plasticity and the
formation and strengthening of long-term synaptic bonds should be explored in
more detail to fully answer this question.
Consciousness as a
Standing Field Formation
While Signal Theory focuses
primarily on the movement of electrical current through the neocortex, it should
also be noted that any electrical field passing through a channel also creates
an electromagnetic (EM) field, and the same is true for the electrical pulses
in the brain. If Signal Theory is extrapolated to its fullest potential, one
would expect to see observable results in the EM field which represents the
process of consciousness moving from one optimized state to another. If this is
so, then distinct states of consciousness can also be modeled in terms of the
field properties which accompany signal interaction through the neural
circuitry.
Isomorphs of Iterative Feedback Processes and Psychedelic Action
- Mediation and
Biofeedback
- Video Feedback
- Audio Looping and
Filtering Popularized in Acid House and Trance Music
- Tides and Weather Systems
- Fractals
- Genetic Expression
- Consciousness
The Future of Signal Theory
If the base principles of
Signal Theory are true, then we will have a new model for describing targeted
states of consciousness in terms empirical signal properties. Everything I am
asserting here can be tested both experientially and objectively via cranial
monitoring and scanning technology, and I would hope that psychedelic and
consciousness research in the next few decades could either produce evidence to
corroborate this theory or at least find one which more accurately demonstrates
how simple psychedelic action in the cortex can generate the wide varieties of
unique mind states and perceptual experiences associated with classic
psychedelic phenomena.
The ramifications of Signal
Theory are currently being explored in the text Psychedelic Information Theory, by James Kent
(jamesk@tripzine.com). For more information, please visit http://tripzine.com/pit

References
·
Best, Ben; An Overview of Neural Networks;
[link] Good overview
of how to model complex neural networks in a computational environment.
·
BilZ0r. "Neuropharmacology
of Hallucinogens : a brief introduction". Erowid.org,
v1 Feb 2004. [link]
Good overview of psychedelic 5-HT2A receptor interaction.
·
Hobson,
J. Allan; The Dream Drugstore: Chemically Altered State
of
·
J. Hunter, Milton, Thomas, Cowan; Resonance
Effect for Neural Spike Time Reliability; The Journal of Neurophysiology Vol. 80 No. 3 September 1998, pp.
1427-1438. [abstract]
·
LeDoux, Joseph; Synaptic Self: How Our Brains Become Who We
Are. Viking Press,
·
LeDoux, Joseph; The Emotional Brain: The Mysterious
Underpinnings of Emotional Life. Touchstone Press,
·
Longtin , André; Autonomous stochastic resonance in bursting
neurons; Phys. Rev. E 55, 868-876 (1997).
[abstract]
This article is primarily about stochastic resonance, but contains formulas
for measuring neural spike phase locking in response to resonant sub-threshold
oscillations. (need to buy full article to read).
·
Lumer, Edelman, Tononi;
Neural dynamics in a model of the thalamocortical system.
II. The role of neural synchrony tested through perturbations of spike timing; Cerebral Cortex, Vol
7, 228-236, Copyright © 1997 by Oxford University Press. [pdf] Interesting
article detailing methods for modeling synchronous oscillations and feedback
loops within thalamocortical and corticocordical
layers of sensory processing.
·
Mato, G.; Stochastic
resonance using noise generated by a neural network; Physical Review E;
Vol. 59 No. 3 March 1999; pp. 3339-3343. [pdf] Mato’s article describes a way to mathematically
model synchrony and resonance in complex neural networks.
Revision History
1.04 – 1/13/2006
·
Rewrote opening to remove entheogenic discourse
altogether. Will move to a different essay.
·
Added section, “Foundations of Consciousness” to
give crash overview of cognitive models
·
Added brain illustration to “Schematic Model”
section.
·
Update References.
1.02 – 11/12/2005
·
Added References
1.01 - 11/10/2005
·
Per
suggestion by Jon Hannah, rewrote opening and closing discourse to remove
loaded terminology.
·
Added
more detail to intro text to frame theory in both brain-based and spirit-based
models.
·
Added
Resonance (rs) as its own primary factor in
calculating synchrony of firing patterns and recursion intensity within a given
circuit.
·
Added
dopamine hypothesis of neural synchrony to questions.
1.00 - 11/07/2005
·
Public
release of Signal Theory Overview.