Neurotransmitters & Neuromodulators: The Psychedelic Connection Neurotransmitters & Neuromodulators: The Psychedelic Connection
James KentChapter 08: Psychedelic Information Theory Although neural impulses fire down the cell membrane in a charged electrical wave, the way these impulses pass from one neuron on to the next is not through electrical charge but through a spray of chemical messengers called neurotransmitters. Now anyone who knows anything about psychedelics knows that the vast majority of psychedelic compounds are structurally similar to if not nearly identical to the neurotransmitters we find naturally occurring in our own brains. Some of the most basic psychedelics (such as DMT) can be organically synthesized by raw materials in our own brains, and the production of endogenous psychedelics (meaning synthesized organically within the body) has been proposed as a possible explanation for a variety of mental phenomena including dreaming, daydreaming, hearing voices, epiphanies, acute schizophrenia, alien abductions, seeing God, and falling in love, just to name a few.
Because psychedelics are so similar to our everyday neurotransmitters it is very tempting to assume that psychedelics act like "false" or "extra" neurotransmitters that float around in the neural mix, sticking to random receptors and basically messing things up in the delicate brain soup. Although this is a gross description it is not entirely inaccurate, but it still falls extremely short of a full working explanation for the entirety of psychedelic action and experience. The prevailing sentiment that psychedelics "mess up your head" offers a very narrow view into the wide range of effects that these substances are known to produce. While psychedelics can certainly mess up your head they can also un-mess your head and make your brain, mind, and body do all sorts of things you never imagined they could do. How does that all come about from a few fake neurotransmitters floating around in the neural soup? The theory of action is there, but just because a substance is similar in structure to another neurotransmitter does not explain the variety and intensity of the state it produces, it only offers tantalizing hints. So let's take a closer look.
The Hallucinogenic Amines
Figure 3: Structurally Similar
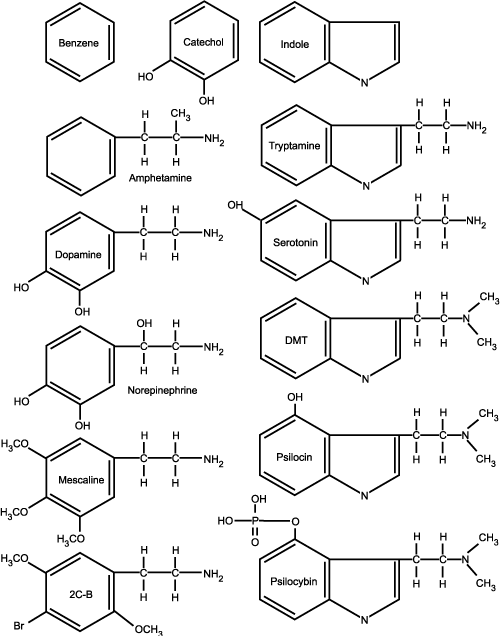
Psychedelic amines look very much like our endogenous neurotransmitters (serotonin, norepinephrine, and dopamine). The 6-carbon benzene ring is the starting point for the amphetamines, the phenethylamines (phenol being any benzene structure with at least one hydroxyl group attached to the ring), and the catecholamines (catechol being a benzene ring with two hydroxyl groups attached to the ring). The basic indole ring structure is a 6-carbon benzene ring with an extra 5-sided ring containing 4 carbons and a single nitrogen, and is the starting point for all of the tryptamines, also knows as indoleamines or indolealkylamines. The actual amine structure of each molecule is the nitrogen group (or tail) that hangs off the backbone of each type of chemical. By changing the shape of the amine tail or by making additions or substitutions to the root ring of each chemical type, you can produce an endless array of "designer" amine derivatives, which may or may not be psychoactive.
[More Images]
|
|
If we take a close look at the neurotransmitter picture we see that the endogenous neurotransmitters most closely related to the classic psychedelics are serotonin, adrenaline (or epinephrine), norepinephrine, and dopamine, and all of these chemicals are classified as amines, meaning that they have a nitrogen-containing amino group (NH2) hanging off the tail of the root structure. This nitrogen structure is the key element in any amino acid, essentially carrying the energy needed for metabolic processes in the body to expend energy and do work. Since these transmitter chemicals have only one nitrogen group they are called monoamines, and are referred to as the essential messengers of the aminergic neuromodulatory system. So when you see amines on the end of any word that means nitrogen, and little crystal structures that carry nitrogen atoms from one place to the next. Endogenous monoamine structures are typically kept out of the brain by the blood-brain-barrier because they carry an electrical charge, but psychedelic molecules are neutrally charged and slip right through no problem, and when these amine crystals surge through the blood-brain barrier they brush against receptor sites dangling off the local neural fauna. If the receptors are a good fit then the crystals get stuck, much like billions of tiny kites getting stuck in billions of tiny trees (or patches of nano-anemones, as the case may be). This receptor activity is where psychedelic action begins.
The Indoleamines, Serotonin, & The Psychoactive Tryptamines
The most popular of the neurotransmitters is serotonin (or 5-HT, 5-hydroxytryptamine). Everyone loves serotonin. It is fundamental to just about all of the basic brain functions, linked to mood, depression, contentment, anxiety, sleep, appetite, and instrumental in regulating the involuntary smooth muscles that control blood pressure and digestive functions within the body. serotonin is an indoleamine and a variant of tryptamine, which is the most basic of all the indoleamines and the structural starting point for DMT (N,N-Dimethyltryptamine), 5-MeO-DMT, Psilocin, Psilocybin, DPT, AMT, and so forth. LSD is also a tryptamine, but it is larger and more complex than the basic tryptamines mentioned above, and is in many ways structurally unique. It seems obvious that hallucinogenic tryptamines most likely exert their influence on the mind via interaction with the various seratonergic systems, an idea we will explore in great detail throughout this text.
The Catecholamines, Dopamine, Norepinephrine, & Psychoactive Phenethylamines
The second favorite neurotransmitter is obviously dopamine because it has the word "dope" right in it's name. It is synthesized from l-dopa (which sounds like the name of a Mexican superhero, !el dopa!), and is considered to be instrumental in modulating fine motor control and attention. Dopamine imbalances are linked both to Parkinson's disease at one end (extreme interruption of dopamine modulation, thus resulting in shaking and loss of motor control), and psychosis at the other end (or extreme dopamine modulation, invoking manic impulsivity, paranoia, and audio hallucinations). The role of dopamine interruption is obviously relevant to psychedelic activity, as psychedelics have been known to cause both shaking and paralysis as well as psychosis.
The phenethylamine group of psychedelics (such as mescaline, 2-CB, MDA, MDMA, etc.), are more structurally similar to dopamine (3-hydroxytyramine), epinephrine (adrenaline), and norepinephrine (noradrenaline), which are all monoamines and sometimes referred to as catecholamines. Epinephrine and norepinephrine are often referred to as "stress hormones" which kick-start the body's energy production in response to stress and danger. The phenethylamines and catecholamines all use the simpler six-carbon benzene ring backbone with at least one amine (nitrogen) group hanging off the root structure. The simplest form of this molecule is called Phenethylamine, and is structurally similar to amphetamine. In very general terms, the tryptamine psychedelics are thought to be more visually hallucinogenic, "trippy," and "far out," while the phenethylamines are said to be more "empathogenic" and "sensual." These descriptions are very broad and of course each psychedelic can produce a range of experience which crosses these vague boundaries, but in general terms this is the popular perceptual distinction made between the two major classes of psychedelics, and we will be discussing the precise effects of various substances in a later section.
Neuromodulators, Mood, & Global Brain States
A lot of recent pharmaceutical research into the treatment for anxiety and depression has focused on how to manipulate the available levels of serotonin in the brain. While some basic corollaries have been found between the amines and mood, nobody really knows to what extent serotonin and dopamine fully affect mood, function, and behavior. What we do know is that serotonin, dopamine, and the other amines don't actually cause neurons to fire, but they do modulate the firing of neurons, which means that they can depolarize or polarize neurons and make them more likely or less likely to fire. In general, aminergic modulators like serotonin and dopamine are projected into higher areas of the brain from the brainstem or middle brain, meaning that they are very ancient and primal signaling mechanisms that are now in charge of controlling large areas of our brain all at once. In effect, neuromodulators are there to fine tune the neural response to incoming stimulus (slow it down, speed it up, shut it off, etc.) and to keep the various disparate brain circuits all functioning smoothly and in unison. And though there is some feedback response from the higher brain which affects the flow of endogenous amines coming from the brainstem, these neuromodulators mostly tend to produce a bottom-up effect, which means they are switched automatically and unconsciously as directed by the brainstem. In this way, the oldest and most basic portion of our brain can regulate cellular messaging functions throughout the entire body without us even thinking about it, and the effect of aminergic modulators on the neuron are tonic, which means their effects are sticky and last over time.
Generally serotonin is thought to have a polarizing effect on neurons, making them less likely to fire and thus having an overall relaxing effect on the brain. This is why many depression and anxiety remedies focus on increasing the supply of serotonin in the brain through one mechanism or another. Now if we take the analogy of "fake" neurotransmitters and apply it to DMT, we would expect that because DMT is so structurally similar to serotonin that DMT should also have a calming, anti-anxiety affect on the brain. Wrong! Nothing could be further from the truth! Structurally similar drugs may act on the same receptors as normal transmitters, but in addition to acting as mimics (mimicking the effect of an endogenous chemical) they can also act as agonists (meaning that they aid in or amplify the flow of chemical signal hitting the receptor) or antagonists (meaning they slow down or block the flow of signal at the receptor). Since DMT sticks to the same receptor sites as serotonin (5-HT2A and 5-HT2C receptors), but does not produce a relaxing seratonergic effect, it would be logical to assume they are most likely acting as 5HT2 antagonists, which means that they are blocking the calming effect of serotonin, thus depolarizing neurons and making them more excitable.
This antagonist theory would be a logical conclusion, but the answer is not always so easy with pharmacology, especially with psychedelic pharmacology. After a lot of research, scrutiny, and debate in this area, the reality seems to be that psychedelics can act in both partial agonist or antagonist capacities depending on the specific drug, the dosage range of the drug taken, the specific receptor subtype it is acting on, and the specific stage of metabolic breakdown the molecule may be in. So let's own up to the fact right now that this is a complex, dynamic equation that has a range of effects over the duration of the interaction, and that reducing the psychedelic experience to one specific interaction at one specific receptor subtype (while tempting), is probably the least likely of correct answers. We are looking at a broader range of interaction here. A lot of stuff is going on.
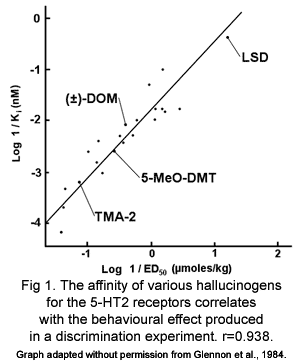
We will come back to the notion of dynamic receptor interaction in detail throughout the course of this text, but it is important to know that while psychedelics may stick to many of the same receptor sites as the endogenous amines, the type and duration of effect at each various receptor subtype is different for each and every psychedelic compound. The strength and duration of the bonds that a ligand (such as a drug, or neurotransmitter) forms with each receptor site is referred to as receptor affinity. The higher the affinity the stronger and longer the ligand bonds to the cellular structure. The groundbreaking work of Dr. Dave Nichols, co-founder of the Heffter Research Institute and Professor of Medicinal Chemistry and Pharmacology in the School of Pharmacy and Pharmaceutical Sciences at Purdue University, has shown fairly conclusively that higher receptor affinity equals higher potency of the psychedelic compound, and the conformational shape of the "tail" of each psychedelic amine plays a big role in both how long it takes to metabolize and how attractive it will be to each receptor subtype. For instance, the "tail" of LSD is different from other tryptamines, it is long, complex, and connects back to the benzene ring lashing it in place. The amine tail on un-leashed molecules are flexible, they can wave around a bit. But the size and conformational rigidity of the LSD may play a role in metabolic rate and receptor affinity, thus increasing the action and varieties of action at various receptors over time. By using this same "tail lashing" technique with new designer amines, Dr. Nichols has shown a marked increase in drug potency (in animal studies) to support this simple theory conformational potency.
If we take that knowledge for what it is worth, it can be assumed that the unique structure of LSD's elaborate amine structure is what makes it so potent; giving it a high affinity across a wider range of receptor types (5-HT1, 5-HT2, D1, D2), making it more difficult to metabolize, and giving it a broad range of effects over a longer duration. DMT, on the other hand, has a very small tail, sticks to a smaller number of receptor types (5-HT2, TA), and is metabolized very quickly. However, DMT's strong affinity to the 5-HT2 receptor type is what makes it such a potent visual hallucinogen, and it is very good at what it does, much better than LSD even. And what the rainforest shamen discovered about DMT way before Western pharmacologists, is that when taken with a monoamine-oxidase inhibitor (MAOi), the enzymes which metabolize DMT (and other monoamines, such as LSD and serotonin and dopamine, etc.) are blocked, and thus the effects of DMT (and other monoamines) last much longer. This is the secret of the ayahuasca brew of South America: the DMT from the psychotria viridis leaf becomes pharmacologically active only when brewed with the harmaline containing yage vine, which acts as an MAO inhibitor. So, there's one scientifically useful fact we've found in the shaman's old bag of tricks, but surely not the last!
Glutamate and GABA: Neural Stop and Go
Figure 5: Amino Acid Transmitters
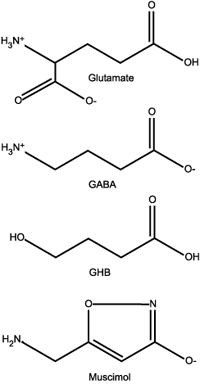
Glutamate and GABA are the workhorse chemicals of synaptic transmission, the go/no-go functionaries that keep neurons firing and keep them in check. Both GHB and muscimol (the active ingredient in amanita muscaria mushrooms) act on the GABAergic systems.
|
|
So far we have talked about the aminergic neuromodulators, but as said before, these are not the chemicals that make neurons fire, they only modulate or fine tune the rate of neural firing. The neurotransmitter that actually sends the "fire" signal from one neuron to the next is glutamate. When a glutamate molecule hits a glutamate receptor it is like a little key which opens a selective sodium ion (Na+) channel to coax the neural chain reaction further along. If a single neuron is hit with numerous incoming glutamate transmissions all at once it will depolarize and then fire. But the entire neural firing model is not quite that simple. Just as glutamate opens a selective channel for positively charged sodium ions, the neurotransmitter GABA opens a selective channel in the GABA receptor to let in negatively charged chloride ions (Cl-), which in turn dampen the effect of any incoming positive ions. Thus, glutamate is a promoter of neural firing and GABA is an inhibitor. While glutamate coaxes the charge along, GABA acts as a buffer in the processing system to keep neurons from firing unchecked at the slightest impulse. Without the balancing effect of GABA, over saturation of glutamate in the brain would cause neurons to fire themselves to oblivion, literally bursting at the seams from over stimulation. This kind of cell death is common during the brain's frantic reaction to a stroke, for instance, when glutamate from ruptured neural cells pours out and causes a chain reaction of toxic over-stimulation, or excitotoxicity, in nearby neurons.
So glutamate and GABA act directly at the receptor to control voltage across the membrane, but the monoamine neurotransmitters like serotonin, dopamine, epinephrine, and norepinephrine act as modulators to facilitate or inhibit the actions of both glutamate and GABA on the cell membrane. Also, unlike glutamate and GABA, which have fast-acting transactions at their receptor sights, the modulating neurotransmitters have slower-acting, or tonic, effects on selective permeability which can either increase or decrease the effect of GABA or glutamate at the individual receptor site over many repeated firings.
It is worth mentioning again that while glutamate and GABA are produced in all neurons in the brain, the monoamines like serotonin and dopamine are only produced by cells in the brainstem. Although located in a primitive part of the brain, the axons from these cells reach outward to many areas of the cortex, and are able to affect the thalamus (primary sensory processing station), amygdala (fear and survival instinct), hypothalamus (body temperature, hunger, thirst, sexual activity, hormonal and emotional response), hippocampus (learning and memory), and neocortex (speech, sensory processing, logic processing). Through the diffuse projection of these neuromodulators, the impulses from the primitive brainstem can have a global effect on the entire brain all at once, thus causing the entire brain organ to speed up, slow down, amplify, or filter signals as it sees fit. GABA and glutamate are the switches which maintain the balance of neural firing at the cellular level, but monoamines like dopamine and serotonin exert greater control at a organism level, causing synchronous global shifts in brainwave activity and mood.
And if you've already noticed that GABA sounds (and looks) like GHB, then yes, you have guessed that GHB works at some of the same receptor sites as GABA. So does alcohol, ketamine, DXM, PCP, valium, and host of other narcotics and dissociatives, but we'll get to that later.
Acetylcholine, the Cholinergic System, Dreaming, and AIM
Figure 6: Cholinergic Modulators
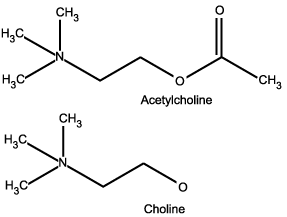
Acetylcholine is the major neurotransmitter in the cholinergic modulatory system within the central nervous system.
|
|
While the majority of the discussion on psychedelic receptor affinity and neurochemical interaction focuses on the aminergic systems, it is important to point out that all of these receptor specific signaling systems rest in a delicate balance with all the other systems within the brain. If activity in one system goes up or down there will undoubtedly follow a chain reaction of corollary or inverse reactions in other systems to mediate the balance of activity. For example, an increase in serotonin supply can also trigger an increase in dopamine supply, and vice versa. But perhaps the most dramatic dynamic in the brain is the chemical switching gear which mediates the shift between waking and sleeping, states which are commonly considered to be two wholly separate realms of consciousness. But are they really? And if so, where is the actual handoff between the two?
This was not an easy question for science to crack, but in recent years it has become clear that sleep-onset, sleeping, and dreaming are chemically mediated in the brain by an increase in acetylcholine and cholinergic activity, with an inversely proportional decrease in serotonin, norepinephrine, and aminergic activity. The interplay between these two systems acts like a tide between waking (aminergic) and sleeping (cholinergic) worlds; when one washes in, the other washes out, and vice versa. So one can assume that drugs which increase aminergic activity (like antidepressants) should subsequently decrease cholinergic activity (dreaming), and in the case of antidepressants and dreaming this is true: when aminergic modulation (serotonin supply) goes up, cholinergic activity (dreaming) goes down. And it has also been shown that prolonged suppression of cholinergic activity and REM sleep (such as sleep deprivation or prolonged use of amphetamines) can lead to psychotic episodes, which may be crudely defined as bursts of delusional or psychotic dream activity erupting spontaneously into waking states.
The delicate interplay between the aminergic and cholinergic systems in modulating waking, dreaming, and other altered states of consciousness has been most succinctly expressed by J. Allan Hobson in the landmark text The Dream Drugstore: Chemically Altered States of Consciousness, which calls upon his years of research and practice in the fields of Dream Research, Psychiatry, and Neurophysiology to create a single unified theory of consciousness. Hobson hypothesizes that the interplay between aminergic and cholinergic systems is what mediates not only waking and dreaming states, but all the hypnagogic, psychotic, delusional, maniacal, and altered states which can occur when these two systems become unbalanced and overlap with one another. To map specific states of consciousness, Hobson created a three-dimensional array that would plot following variables at any given moment: A) Activation of the brain (high energy or low energy), I) Input of signal (internal vs. external transmission, circuits open vs. closed), and M) Modulation (aminergic vs. cholinergic). By plotting these three coordinates (AIM) Hobson has provided a way to generalize a neurochemical snapshot of the brain in a graphic context.
Figure 7: AIM State Spaces
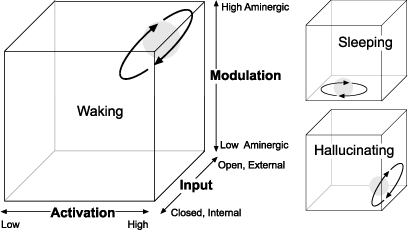
Hobson's AIM model maps levels of brain Activation, the gating of signal Input from external to internal sources, and changes in Modulation from aminergic (waking) to cholinergic (sleeping/dreaming) systems. Under the influence of hallucinogens, Hobson's model predicts high activation, a disruption in aminergic functioning, and a selective gating of input from external to internal sources. Hence, a marginal "waking dream state" emerges.
[More Info]
|
|
Hobson's AIM model hypothesizes that hallucinogenic amines work by selectively shutting down the aminergically modulated (waking) system and nudging the balance of the cholinergic system to the fore, thus a "waking dream" erupts. In this state, the rational and contextual reasoning areas of the brain which are modulated by amines (serotonin, norepinephrine) are dampened, while the more emotionally driven and imaginative acetylcholine-modulated parts of the brain are turned up. Hobson's theory is so elegant and well researched it is hard to find fault with any of his assumptions, and I would highly recommend The Dream Drugstore to anyone who wishes to explore the neurodynamics of the AIM model in more detail.
However, Hobson admits that he has never taken psychedelics, and bases most of his theory on his own dream and lucid dream research, plus his expert knowledge of psychosis and delusional states. While his theory provides a good model for examining psychotic states in general, I thought perhaps he might be missing something about the psychedelic state by oversimplifying it as a kind of semi-lucid dream-state or waking psychosis. I have little doubt that some, if not all, psychedelic action is mediated exactly as Hobson hypothesizes, but he misses one key point about psychedelic effect in his effort to lump it in with lucid-dreaming and psychotic states, and that is this: The psychedelic "trip" is a totally dynamic state-space and may wander all over the map between lucidity, psychosis, lucid dreaming, dreaming, hyper-awareness, delusional states, mystical states, mania, delirium, etc. In a sense, the one thing missing from Hobson's model is the will of the individual and the context of the trip which actually snaps the fully psychedelic state into focus, instead of having it trail off into delusional or psychotic ideation as Hobson's model would indicate.
While being so intently focused on lucid-dream states, Hobson seemed to miss a critical point about psychedelics that I will define thusly: psychedelics are to the entirety consciousness what lucid dreaming is to dreaming. In other words, the word 'psychedelic' does not necessarily conform to one specific spectrum of consciousness, it encompasses the totality of targeted effect amplification across the entire spectrum of consciousness. In the lucid dream state, all of the possibilities of the dream world become within your grasp, but the grasp is slippery and fades quickly. In the psychedelic state, all the possibilities of the waking and dreaming worlds become within your grasp, the grasp is still slippery but it does not fade quickly, and given proper training it can even be wrangled. And as Hobson discovered with lucid-dreaming, you get better with practice, and the same is true with psychedelics; you eventually learn your way around the territory and can do "whatever you want" as long as you can think it up. Consider that application shifted from the lucid-dreaming world to the lucid-waking world. Whoa! Heavy man!
In essence, the difference between what Hobson and I would have to say about psychedelics is very little, but I would argue that psychedelic action is not limited to any one particular plot in the AIM space, but could easily move through all of the spaces within that map in a single trip. Or, just as easily, the psychedelic state could park right in one spot and stay there for almost the entire duration of a trip, but that destination may be different for different people, and appear at different spots on the AIM space depending on dose and context. As I said, people take psychedelics for a lot of complex reasons, and the things we expect from them are equally complex. The lucid-dream or "waking dream" state is just one of the more obvious psychedelic sates, but I would say it differs greatly from the truly entheogenic or other "lucid-waking" psychedelic states which are even more subtle and complex to unravel than the simple tidal interplay between the cholinergic and aminergic systems would initially indicate. But Hobson's work on the boundaries of waking, sleeping, and waking psychosis no doubt are an excellent jumping-off point for our quest, and we will come back to Hobson's unified theory of consciousness many times within the course of this text.
Neural Circuitry: Excitation, Inhibition, and Autostimulation
Now that we've discussed the dramatic effects that neuromodulators can have on mood, global brain states, and signal flow, we now get into even more complicated territory: the inter-neural mediation of signal firing. For every projection neuron — a neuron which signals to another neuron down its axonal branches — there are a series of interneurons which provide feedback from one part of the neuron to another, thus giving each neuron the ability to act as it's own feedback amplifier (or filter) for every signal that it processes. The action or inaction of these interneurons is generally dependent on the presence of the modulating chemicals we just discussed, each having either a dampening or promoting effect on the general sensitivity of each neuron. In the presence of the psychedelic, specific neurons may become hypersensitive to feedback stimulation, thus causing a state of neural autostimulation in some areas of the brain. As we will discuss in a moment, autostimulation and feedback promotion mediated through interneurons may well account for many of the "over the top" temporal, auditory, visual, and spatial distortions perceived within the psychedelic state.
When a neuron is so highly sensitized that it will fire at the slightest provocation, this is referred to as a state of feed-forward excitation or elicited excitation, in which the interneurons increase the amount of feedback stimulus each neuron receives. In addition to feed-forward excitation, a neuron can be in a state of tonic excitation, which is a low-level increase in neural sensitivity over a period time, generally in the presence of a modulating molecule with the proper receptor affinity to achieve that goal. Neural sensitivity promoters can either make the neural membrane more porous to positive ions, less porous to negative ions, or both, causing the charge differential across the neural membrane to temporarily move closer to the action potential (depolarize). When a neuron is excited, or depolarized, it takes less of a stimulus to make it fire, and when it hits feed-forward excitation it can reach a point of autostimulation, in which it's own firing action causes it to re-stimulate itself, causing it to fire again and again until it is spent and must take a break to reset itself. If not given a chance to rest, an extremely hyperstimulated neuron will eventually burn itself out, resulting in cell death. In the case of stroke, it is not the loss of oxygenated blood to the head which causes the most brain damage, it is a flood of glutamate in the brain from ruptured neurons that causes nearby neurons to begin firing unchecked, resulting in cell death by over-excitation, or excitotoxicity.
In certain circumstances a depolarized neuron may begin to fire spontaneously. These involuntary firings may result in a muscle twitch, a nervous tick, or other spontaneous recurring thoughts or sensations. Everybody has these from time to time, a eyelid flicker, a jittery leg, a song repeated internally over and over, etc., and in extreme cases the spontaneous eruption of neural firing can result in a seizure, which is like a storm of unchecked neurons going off all at once. While a muscle twitch, a repetitive ideation, and an epileptic seizure might not seem to be connected at the surface, all three of these behaviors can be traced back to some unchecked autostimulative behavior occurring at the neural level, but in different areas of the brain and at different levels of excitation. In short, a hyperactive neuron or group of neurons is acting out of synchrony with the rest of the brain, throwing localized areas of the normally smooth running order into momentary chaos. And in all cases, extreme versions of these conditions are treated with drugs that inhibit neural firing and keep them from becoming hyperstimulated, and the effective treatment of all these spontaneous conditions comes from targeted neural inhibition of the right kind of neurons.
The reason I bring all this up, of course, is to shine some light on what would happen if the reverse was true, and if targeted neural autostimulation were possible with the introduction of the right kinds of molecules. With the right kind of targeted neurostimulators, we would effectively be able to amplify the performance of any area of the brain — including the rational, mystical, emotional, memory, motor, and/or audio-visual rendering capacities — any time we wanted or needed to. And, of course, any chemical which interferes with the basic neuromodulators (serotonin, dopamine, norepinephrine, acetylcholine) could have such an effect at the neural level, and since psychedelics have been known to act as "Nonspecific Amplifiers" of mental functioning, it is worth looking into the potential "targeted autostimulation" action of psychedelics in more detail within this text.
Neuromodulators & Signal Flow: AIM vs. Signal Theory
Hobson's aminergic/cholinergic AIM model would indicate that the aminergically modulated systems (like the executive self located in the pre-frontal lobes) should be dampened or shut down in a psychedelic trip, while the cholinergically modulated systems (like the thalamus and limbic system) would be stimulated or activated, thus causing the emotional, intuitive (dreaming) brain to come to the fore while the rational, contextualizing (waking) part of the brain goes offline for a little while. This is a reasonable explanation for one specific kind of psychedelic trip, which is the lucid daydream or the 'dream journey', which is typically spent in closed-eye meditation, laying down, somatically overcome with visions, etc. However, not all psychedelic trips are like this, and many are hyper-focused in the real world. Perceptual distortions and peripheral hallucinations are folded into the waking mix, but these kinds of visual distortions are certainly not the same as dream activity, and are most likely being generated in a different way.
The reason I assert this is that I have always been a lucid dreamer, ever since I was a small child, and have always been very aware of that murky layer between awake and asleep where lucid dream activity happens. However, I have also experienced a variety of different psychedelic trips, and while some have certainly induced dream-like activity and dream-like delusion, many have been nothing like dreams. In short, it seems that the paradox of psychedelics is that they can act as stimulants to some parts of the brain and depressants to other parts, or stimulants to many different parts all at once. In truth, much of this paradox can be reduced to simple factors like dosage: a smaller dose of psychedelics is more like an amphetamine, causing the user to be twitchy and hypersensitive to stimulus, while a larger dose is so overwhelming that the user erupts into the somatically overwhelming dream-like delusional state Hobson's model predicts. Other variants can be tied to specific psychedelics which may be more like amphetamines or delirients than the classic psychedelics. But there is also something more going on here.
What I am postulating is that there is something else going on in the psychedelic state besides a variation on the themes of amphetamine psychosis or dream-delusion. What I think is essential to realize about a psychedelic trip is that it can be both of these things, but there is also a happy medium between these two poles, a "sweet spot" where the hyper-activated waking brain and the sensually stimulated dreaming brain can overlap in a moment of cross-synchrony, coupled with the very real effects of autostimulated neural feedback loops and distorted neural transmissions introduced at various levels of sensory signal processing. The end result of an experience like this would not only be magical and dream-like, it would also be a full engagement of all of the brain's systems in one massive multi-threaded parallel contextual sensory processing event; an extreme emotional wringer; a conceptual mind-blower; an explosive lid-flipper; and a full body-mind-and-soul workout from the top down, the bottom up, and the inside-out. And this is precisely what the most sought after psychedelic experiences are like, which is why I will continue to return to this notion of sensory feedback loops and cross-lobe experiential amplification from the neural level on up throughout the course of this work.
RNA Editing: Secondary & Tertiary Messengers
Figure 8: 5-HT2 Receptor RNA Editing
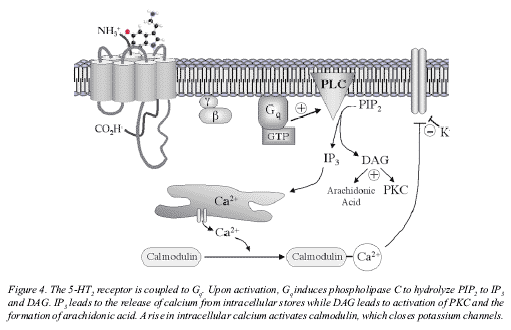
Not quite as diabolical as it sounds, RNA editing across the neural membrane is achieved by a torsion in receptor rungs as the receptor clings to the ligand. This torsion causes the intracellular RNA loops which code for specific proteins to bend, disrupting internal interactions with intracellular RNA transcription enzymes.
[More Info] [More Images]
|
|
Of course, there is much about the neuron and neurotransmitters I have left out and cannot possibly cover within the scope of this text, but what we are primarily interested in here is the flow of information through the neural network, and we have at least covered the basics. There are many ways in which a neuron can be depolarized or polarized, but the most common is through direct neurochemical action at the receptors, and this is how we assume psychedelics elicit the majority of their effects. However, there is some recent evidence that shows LSD may have a short-term conformational effect on the 5HT2 receptor's RNA complex, which in turn affects the production of secondary messengers on the inside of the neural membrane. In other words, the action of LSD hitting the receptor changes the shape of the RNA strand, keeping it from reacting properly with transcription enzymes that code intra-neural signaling proteins, or secondary messengers, in the cytoplasm of the cell. These secondary messengers interact with tertiary messengers at the cell's nucleus and can affect a wide array of biological processes, including a change in the intracellular charge and/or a disruption to the various catabolic and metabolic pathways within the neuron.
If LSD (and other psychedelics) do affect the transcription of messengers on the inside of the cell membrane at the 5HT2 receptor (the one primarily implicated in hallucinogenic affect), it is possible that psychedelic action may be more complex than we originally thought. Weather or not the RNA editing effect on the coding and production of secondary messengers has anything to do with a shift in consciousness is speculative territory. However, for the purposes of this discussion (which is about signal flow), even simple affects on secondary messengers within the neural membrane may result in many significant outcomes: 1) The production of essential messenger chemicals within the cell body causes a reversible, short-term imbalance (shortage or oversupply) of specific intracellular materials, which could affect the coding of memory, the speed of neural firing, and various other intracellular processes. 2) The neuron may be either more polarized or less polarized as a result of the new chemical balance, and is thus either less likely or more likely or to fire at the introduction of any specific stimulus. 3) The supply of endogenous signaling neurotransmitters produced by the cell's nucleus may be increased or decreased, thus increasing or decreasing the strength of each signal repetition at the synaptic level. 4) The disruption in the balance of specific messenger types could slow down or speed up intracellular processes which are essential to resetting and/or conditioning the neuron to fire, thus causing signal delay or conversely signal distortion.
Now, the idea of LSD acting in an editing capacity on the transcription mechanisms of your neural hardware is pretty heavy stuff, but it is not an uncommon neural event. Another way of thinking about it is that the unusual shape of LSD causes the receptor to bend in unusual ways to get the best grip on it, and in doing so the rungs of RNA messaging codons gets a little bent out of shape, so the transcription enzymes can't stick to the bent rungs of genetic code as well as they typically should, and others may fit a little better in their place. Again, this may be a very elegant and detailed way of saying, "Psychedelics mess things up in your head," but it is important we look at this particular aspect of RNA editing closely to see the many ways in which signal can be disrupted at the neural level.
Now I'm sure there are some of you out there who might be thinking, "Aha! There is another possible way RNA editing at the 5HT2 receptor site might alter consciousness: LSD (and other psychedelics) may actually send a secret code to the cell's nucleus, thus unlocking a hidden feature of genetic memory that makes you trip!" Okay, before we go off into science fiction territory it is important to point out that secret codes (messaging chemicals) and genetic memory (DNA) are the basics of all cellular function and communication, and "hidden features" are just reactions we haven't seen under a microscope yet. However, saying that psychedelics are a secret code to unlocking hidden features of the mind implies that a programmer put those features there and intended us to eventually discover them, perhaps when we were smart enough to find the proper key. This line of reasoning comes very close to presupposing a God or "master programmer" that has built this feature into human consciousness, and it must also embrace something as far fetched as a built-in "genetic memory" that can be accessed and unspooled like a film-reel at anytime you ingest the right chemicals. Needless to say the possibility of a genetic memory raises more questions than it answers, and I will talk a little more about genetic memory in various places throughout this text. But however far out this idea may be, I cannot tell you the number of times I have had people tell me they were absolutely sure that this is what is really happening in the psychedelic trip, and they couldn't believe I would ever doubt it, so there you go.
Neural Plasticity
Although the issue has been debated for some years, the most recent scientific evidence shows that axons of active neurons are always branching, growing and creating new connections to facilitate learning and development as we grow. While the macrostructure of the brain is fairly rigid, the individual circuits and synaptic connections within a particular organ or region of the brain can vary from individual to individual. Like the toning and bulking of muscle mass, neural connectivity, developmental growth, and relative plasticity are based on the "use it or lose it" principle. The more a neuron or circuit of neurons is used and exercised, the stronger, faster, and denser its connections with other neurons becomes. Under this principle, little-used portions of the brain can suddenly become very robust when in the process of learning a new skill, and robust portions of the brain can deteriorate if they go unused for a long period of time.
While I do not want to out-and-out implicate real-time neural plasticity as a culprit in the origin of content generated within the psychedelic experience, some consideration has to be given to the fact that targeted cross-activation of certain areas of the brain can lead to permanent affinity in the firing patterns between those two areas. What this means is that previously unrelated areas of the brain, such as those that hold the memory of "fish" and those that hold the memory of "cheese" can become virtually fused through repeated cross-activation into a larger concept known as "fish-cheese" which then becomes a new neural connection in your brain, which in turn creates new information. Each time you think of "fish-cheese" your are calling upon the new connection you have made between these two previously distinct areas of your brain. And one of the commonly reported effects of psychedelics is this surreal cross-breeding of novel thought within the hyper-stimulated brain. While it is not clear at what point an actual lasting neural bridge is created in the fusing of new concepts, it is the idea that mind is plastic and that new connections between previously unrelated areas of the brain are being created all the time that I want to impress here.
Toxicity and Adverse Cellular Reaction
So now we know that each neuron is both a little switching station, motor, and factory for the neurochemical flips and switches that mediate our very thought. It is also alive, living, growing. We also know that disruption of any one of these little flips, switches, pumps, channels, signaling chemicals, etc. can result in erratic functioning of the entire neuron, and thus erratic functioning of the entire brain. However, since psychedelics do not destroy neurons, it is safe to say that any effects they have on the neuron are entirely reversible, and do not lead to any cellular disruptions extreme enough to cause cell death. There are exceptions to this statement, of course. Ibogaine (from the African Iboga shrub) has been shown to be toxic to Purkinje cells in the cerebellum in large doses, though the effects of this cell death on behavior are not readily apparent. And there is the infamous MPTP - the synthetic heroin that causes dopaminergic cell death in the substantia nigra and sudden onset Parkinson's Disease - that accidentally hit the streets when underground batches of MPPP (another synthetic heroin that is not toxic) went wrong. But for the most part the psychedelics discussed in this text, when taken in the appropriate dose range, do not cause cell death in any large-scale observable way, at least none that has been adequately measured and reproduced.
However, given that temporal lobe epileptic seizures can cause cell death as well as elicit very strong mystical reactions and/or near death experiences (NDEs), one should not totally ignore the possibility that high doses of certain psychedelics may in fact lead to small seizures in various organs of the brain, resulting in both cell death and/or specific perceptual aberrations along the way. There is limited evidence to show that psychedelics cause seizures or excitotoxicity, and I don't want anyone getting angry at me and saying, "James Kent says psychedelics cause epilepsy and cell death!" because that is not what I am saying. But what I am saying is that it would be foolish to ignore the possibility that psychedelics may in some cases cause localized seizures which may in some cases lead to small-scale excitotoxicity in clusters of neurons, which may or may not lead to temporary or persistent perceptual, emotional, or cognitive effects. But as we have shown time and again with thousands of years of binge-drinking, the human brain can loose clusters of neurons all the time without any noticeable loss of function, so even in these extreme hypothetical cases there is no evidence to show that psychedelics are even near the same realm of neurotoxicity as good old alcohol. Nevertheless, one should always be very conscious of the internal brain state under the influence of psychedelics to minimize any kind of massive disruption, and if you have a history of epilepsy I would say it is definitely contra-indicated for you to be experimenting with psychedelics.
<< Previous | Index | Next >>
Tags : psychedelic
Rating : Teen - Drugs
Posted on: 2005-03-22 00:00:00
|